|
|
|
|||||||||
| |||||||||
|
Калиевые пруды и натриевые океаны. Физические свойства протоклеток и происхождение внутриклеточной среды
В.В. Матвеев, Институт цитологии Российской академии наук,
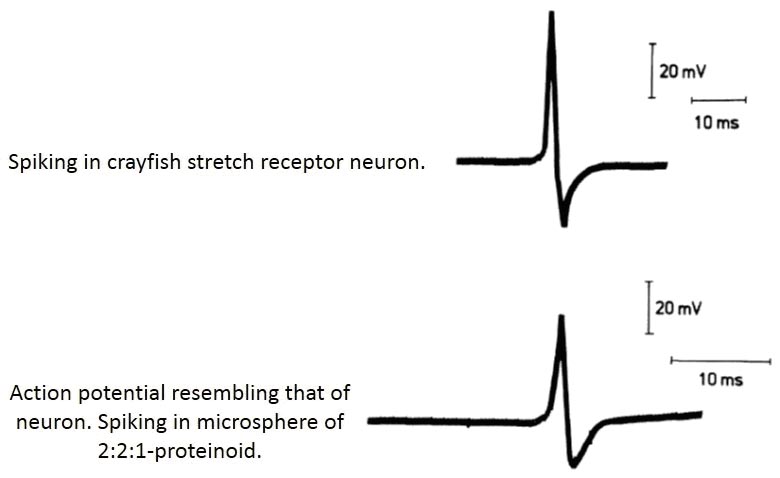
Аннотация . Появившееся недавно предположение о калиевых прудах, как о колыбелях жизни, обогащают палитру идей о возможных условиях предбиологической эволюции на первобытной Земле, но не приближают нас к решению проблемы происхождения жизни в целом. Суть проблемы - в механизме разграничения двух сред, внутриклеточной среды и среды обитания протоклеток. Со времени открытия насосной функции Na, K-АТФазы так и не было найдено модели, которая могла бы рассматриваться в качестве предка современной натриевой помпы, что и вызвало к жизни идею калиевого пруда. Кризис современной методологии настолько глубок, что сталкивается даже с логическими противоречиями при попытке объяснить происхождение первой живой клетки. Одно из них состоит в том, что законы физики запрещают возникновение в термодинамически равновесной системе (пруд) неравновесной подсистемы (протоклетки). Имеющиеся подходы к решению проблемы происхождения жизни полностью игнорируют необходимость детального изучения клеточных моделей. Так, свойства микросфер Сиднея Фокса оказались несовместимыми с общепринятыми представлениями о функционировании живой клетки. Например, мы сталкиваемся со следующим парадоксом: математическая модель потенциала действия Ходжкина-Хаксли оказалась одинаково хорошо совместимой и с нервной клеткой и с микросферами Фокса, также способными генерировать потенциал действия. Цель настоящей статьи - предельно обнажить противоречия, возникающие при попытке объяснить происхождение протоклеток на основе сложившихся представлений о клетках живых. Ставится вопрос о необходимости значительного расширения сравнительных исследований живых клеток и их моделей. Калиевые пруды Излюбленное и научной, и научно-популярной литературой утверждение: ионный состав внутренней среды организма человека и животных, в которую погружены все его клетки, близок к ионному составу морской воды. Это наблюдение попало в литературу уже больше 100 лет назад, когда стало возможным исследование ионного состава биологических жидкостей. Сближение внутренней среды организма с морем напрашивается само собой: и в морской воде, и в плазме крови ионов Na+ на порядок - два больше, чем ионов K+ . Это очевидное сходство и подтолкнуло к мысли о том, что жизнь зародилась в первобытном океане (память о котором хранит с тех пор внутренняя среда организма), а первые клетки отгородились от морской воды плохо проницаемой мембраной для того, чтобы их внутренняя среда стала особой, пригодной для протекания химических и физических процессов, необходимых для жизнедеятельности. Действительно, соотношение рассматриваемых катионов в цитоплазме прямо противоположно их соотношению в морской воде: ионов K + в ней намного больше ионов Na+ . De facto процессы жизнедеятельности возможны лишь в среде, в которой калий доминирует над натрием, поэтому любая теория происхождения жизни должна объяснить, как произошло столь глубокое разграничение между двумя средами: внутриклеточной средой, в которой протекают жизненно важные процессы, и внешней средой, обеспечивающей клетку необходимыми материалами и условиями. При отделении протоклетки от морской среды должен был возникнуть механизм, создающий и поддерживающий ионную асимметрию между первобытной клеткой и ее окружением. Таким механизмом является, как известно, изолирующая мембрана со встроенным в нее молекулярным ионным насосом - Na , K -АТФазой. Если жизнь возникла в морской воде, то происхождение первой клетки неизбежно сводится к происхождению натриевой помпы вместе с поддерживающей ее структурой - липидной мембраной, без которой работа любого насоса не имеет смысла. Получается, что жизнь рождается в условиях, глубоко ей враждебных и вопреки им. Парадоксальность такого хода событий настолько очевидна, что его детали не пытаются даже обсуждать. Трудности, с которыми сталкивается идея происхождения жизни в морской пучине побудили к поиску обходных путей в надежде, что натриевая протопомпа может потребоваться не сразу, а немного погодя, когда в протоклетке возникнет всё необходимое для создания помпы, включая систему ее энергообеспечения. В 2007 году почти одновременно вышли две статьи ( Mulkidjanian and Galperin, 2007; Natochin_2007_eng , Natochin_2007_ru ) в которых высказано предположение, что жизнь возникла не в морской воде, как было принято думать до сих пор, а в небольших водоемах, с нужным для жизни соотношением ионов K + и Na+ : то есть калия в них было больше натрия в той же пропорции, что и в цитоплазме современных клеток. Все этапы химической эволюции, предшествовавшие возникновению жизни, протекали, по мнению этих авторов, именно в таких, калиевых, прудах. Мулкиджанян и Гальперин ( Mulkidjanian and Galperin, 2007;Mulkidjanian et al.,_2012 ) предложили даже «принцип сохранения химического состава», согласно которому в цитоплазме современных клеток законсервирован ряд особенностей калиевых прудов, в число которых, кроме необходимого соотношения калия к натрию, входят, по мнению авторов, и другие металлы, а также некоторые важные для жизни соединения. При таком подходе можно сказать, что калиевые пруды представляли собой, по существу, мегацитоплазму, по крайней мере в том, что касается рассматриваемых катионов. В итоге, образ первобытного океана, текущего по нашим жилам, теперь можно дополнить мириадами калиевых прудов, сохранившихся в составе клеток нашего организма с первобытных времен. Натриевый океан Наточин ( Natochin,_2007_eng , Natochin,_2007_ru ) тоже за сохранение химического состава первобытного калиевого пруда (не выделяя, однако, это положение в принцип сохранения), но в этом первобытном водоеме его интересуют прежде всего физиологически значимые катионы - K + и Na+ . Поскольку калиевая мегацитоплазма не контактирует до поры с натриевым океаном, она, по Наточину, неспособна поддерживать важные жизненные процессы, основанные на физиологическом антагонизме этих катионов. Например, мегаклетка, в образе пруда, неспособна генерировать потенциалы покоя и действия. Отсюда идея Наточина о ключевой роли ионов натрия в происхождении жизни, которая может вспыхнуть только в столкновении калиевого и натриевого миров. Некоторое преимущество калиевого пруда перед морской водой состоит, кроме прочего, и в том, что скорость синтеза пептидов в 3-10 раз выше в среде с ионами калия, чем в среде с той же концентрацией ионов натрия ( Dubina et al.,_2013 ), что можно объяснить разным характером взаимодействия K + и Na + с функциональными группами пептидной связи ( Jockusch et at.,_2001 ). Что касается происхождения таких прудов на первобытной Земле, то одни авторы ( Mulkidjanian and Galperin, 2007; Mulkidjanian et al.,_2012 ) связывают их появление с особыми условиями конденсации водяных паров в регионах с активной вулканической деятельностью, когда в конденсатах оказывалось почему-то больше калия, чем натрия. А Наточин считает, что такие пруды могли возникать и в более спокойной геологической обстановке - при контакте пресной воды с породами, богатыми соединениями калия ( Natochin,_2007_eng , Natochin,_2007_ru ) . Проблемы и суждения Идея калиевого пруда сталкивается со следующими проблемами. 1. Конденсаты водяных паров должны были во многом напоминать дистиллированную воду. Если дистилляция была несовершенной и в конденсате калия оказалось больше натрия, то одного только превышения K + над Na + еще явно недостаточно. Важно и количество солей. 2. Свитек ( Switek, 2012 ), комментируя «принцип сохранения химического состава», считает, что время существования калиевых прудов в неспокойных регионах с активной вулканической и тектонической активностью явно недостаточно для завершения предбиологической эволюции. 3. Являлись ли калиевые пруды колыбелями жизни или они служили только химическими реакторами, в которых накапливались пептиды и другие соединения, необходимые для формирования протоклеток? Свитек убежден, что говорить о возникновении жизни в калиевых прудах нельзя, так как в таких водоемах сравнительно быстро устанавливается диффузионное равновесие всех растворенных веществ. Например, калий в одинаковом количестве присутствовал бы как внутри протоклетки (возникни она в таком месте), так и вне ее, а живая клетка обязана быть термодинамически неравновесной системой. Свитек, хотя и невнятно, по-своему, выражает простую мысль: физические законы запрещают возникновение в термодинамически равновесной системе (пруд) термодинамически неравновесной подсистемы (протоклетки). 4. По Наточину, насколько я могу судить, жизнь может возникнуть только в системе калиевая протоклетка/натриевая среда (т.е. как раз в неравновесной системе). Но в этом случае механизм поддержания неравновесия протоклетки со средой должен был возникнуть загодя, еще в калиевом пруду, в то самое время, когда в таком механизме еще не было практической необходимости! Получается, что в калиевом пруду должны заранее возникнуть липиды и сформировать липидную мембрану. Затем в том же пруду должны образоваться прозапас натриевые помпы и ионные каналы (одни для Na + другие - для K+ ). Наконец, все эти элементы должны каким-то образом собраться вместе, образовав конструкцию, подобную биомембране, и оставаться в ожидании встречи с морской водой то ли через день, то ли через миллион лет. Без всех этих необходимых приготовлений, морской Na+ , катион смерти, беспрепятственно проникнет в клетку и погасит в ней все тлеющие угольки жизни, как только морская вода захлестнет калиевый пруд. Возникает неизбежный вопрос: почему натриевый насос должен был заранее сформироваться еще в калиевом пруду, если в протоклетке, «обитавшей» там, не было избытка Na+ ? Если Мулкиджанян и Гальперин, в нарушение термодинамического законодательства природы, высказываются за возникновение неравновесности (жизни) в равновесном пруду, то по Наточину, протоклетка в калиевом пруду уже загодя готовится к будущей жизни в океане. Ведь, по Наточину, искры жизни появляются только тогда, когда наполненная калием протоклетка оказывается в натриевом окружении. Именно тогда между средой и клеткой возникнет натрий/калиевый градиент, который, как стартер, запустит натриевый насос, заранее для этого приготовленный. Таким образом, идея калиевого пруда приводит нас в к двум неправдоподобным сценариям: если против зарождения жизни непосредственно в калиевом пруду решительно выступает термодинамика, то для зарождения натриевого насоса в условиях, когда в нем нет нужды, необходимо вмешательство Провидения. 5. Свежая, казалось бы, идея калиевого пруда, не оправдав наших надежд, вновь возвращает наше внимание к фундаментальной проблеме: происхождение жизни сводится к происхождению полнофункциональной мембраны, - структуры, способной осуществлять неравновесный процесс по поддержанию особых условий внутриклеточной среды. Калиевый пруд не вносит в эту проблему ничего нового, если не считать новых проблем: что делать с нарушением фундаментальных законов физики, и как быть с теологией, согласно которой, события настоящего определяются потребностями будущего? Микросферам Фокса калиевые пруды не нужны Теперь давайте перейдем от рискованных предположений к установленным фактам. На протяжении последних 90 лет в нашем распоряжении имеются привлекательные клеточные модели, воспроизводящие немало интересных свойств живых клеток (вплоть до демонстрации «ионных каналов», «потенциалов покоя» и «действия»), но и столетия, считай, не хватило для того, чтобы обнаружить модель хотя бы какого-нибудь насоса, из которых нас сейчас интересует натриевая помпа. Ни электрические разряды Миллера, ни аминокислотные сплавы Фокса, ни встраивание в коацерваты уже готовых биомолекул так и не привели к самозарождению первозданного ионного насоса даже в самых благоприятных лабораторных условиях. Мы стали свидетелями своего рода интриги: целые протоклетки, - огромные белковые комплексы, - в лабораториях самозарождаются, а сравнительно небольшие белки-насосы - нет. Отсутствуют даже намеки, куда двигаться дальше в поисках этих ключевых моторов жизни. Не означает ли это, что непреодолимой преградой на этом пути опять встает термодинамика, строго запрещающая желанное чудо -возникновение неравновесности (читай: жизни) в равновесной системе, какой бы она ни была, - рафинированным раствором от «Сигмы», теплым прудом, тихой лагуной или океанической бездной! Ну, а пока науке ничего неизвестно о происхождении ионных насосов, рассмотрим известные свойства микросфер Фокса. На рис. 1 представлены два потенциала действия. Один распространяется по нервному волокну рака, другой - по поверхности протеиноидной микросферы. Из сопоставления двух записей следует, что в обоих случаях изменения электрического потенциала качественно однотипны и количественно близки. Для ясности приведу пример. Силы гравитации на Земле и Луне имеют качественно одну физическую природу, различия носят лишь количественный характер. Наш случай такой же: физическая природа изменения потенциала в аксоне и в микросфере одна и та же (в обоих случая переносчики заряда - ионы), а количественные характеристики различаются куда меньше, чем силы притяжения на Земле и Луне.
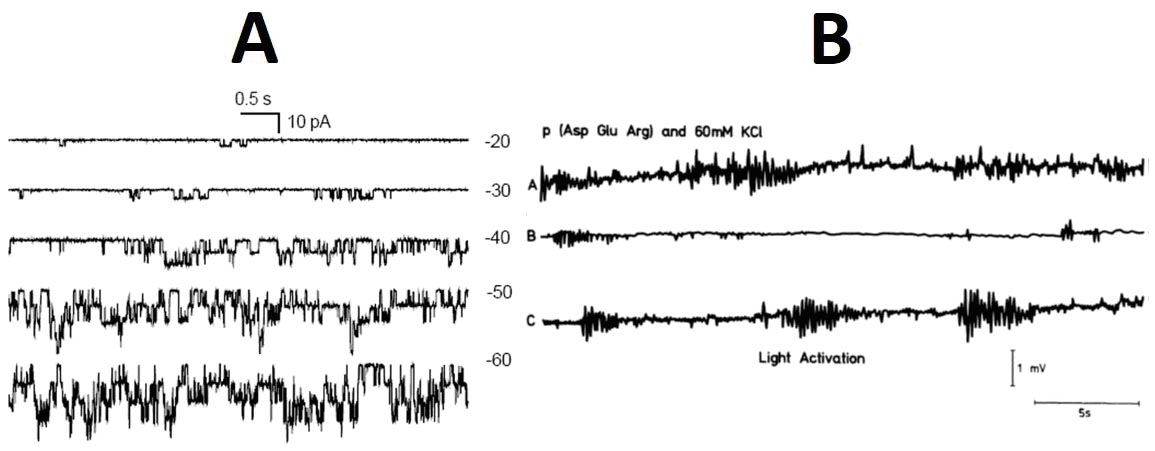
Рис. 1. Потенциал действия, возникающий при стимуляции рецептора растяжения рака (на верху). «Потенциал действия», зарегистрированный у микросферы. По Фоксу (Fox, 1992). Теперь зададимся следующим вопросом: какими структурными элементами должна обладать мембрана аксона для того, чтобы она была в состоянии генерировать потенциал действия? Этих элементов в минимальной возбудимой мембране немного и они хорошо известны: 1) сплошной билипидный слой; 2) калиевые каналы; 3) натриевые каналы; 4) Na , K -насос. Функциональное назначение отдельных деталей этого механизма состоит в следующем: насос создает градиент концентраций K + и Na+ , отчего на мембране возникает разность потенциалов (потенциал покоя); липидная фаза действует как изолятор, подавляющий паразитные токи; ионные каналы служат специфическими электрическими проводниками, необходимыми в данном случае для управления величиной мембранного потенциала. Для восстановления потенциала покоя, после возбуждения, требуется закрыть ионные каналы и включить насос для воссоздания соответствующих концентрационных градиентов. Раскрыть механизм потенциала действия нейрона удалось Ходжкину и Хаксли ( Hodgkin and Huxley, 1952 ), решившим эту проблему с помощью созданной ими математической модели. За эту работу авторы получили Нобелевскую премию в области физиологии и медицины за 1963 год. Из всего разнообразия математических приемов, авторы выбрали математический аппарат, удобный для описания потоков частиц (в данном случае ионов), причем эти потоки разделены в пространстве: одни ионы ( Na+ ) двигаются в мембране по специальным натриевым каналам, а другие ( K+ ) - по калиевым каналам. В целом, как следует из модели Ходжкина-Хаксли, для того, чтобы возбудимость стала возможной, необходима мембрана, переменную и избирательную проницаемость которой описывает эквивалентная электрическая схема, в которой потенциал-зависимые резисторы соответствует ионным каналам, а ёмкостное сопротивление обеспечивает диэлектрическая липидная мембрана. Роль моделей, физических и математических, в науке трудно переоценить. Искусственные мембраны, например, сыграли решающую роль в развитии представлений о роли мембран в жизни клетки. Физические модели атомов и молекул, включая ДНК, математические модели в инженерном деле и астрофизике, все они доказывают, что только соответствующая модель служит если не доказательством, то ключевым свидетельством в пользу предполагаемого механизма явления или структуры объекта. Лучшим доказательством важного места, занимаемого в науке моделями, служит судьба самой модели Ходжкина-Хаксли, на десятилетия определившая не только наши представления о возбудимых мембранах, но и о функционировании мембран вообще. Однако справедливо и обратное: никакой механизм не может считаться доказанным, если не удается найти модель, демонстрирующую, пусть в упрощенном виде, его работоспособность. В данном случае нас, в качестве моделей, интересуют искусственные клетки, представляющие собой камень преткновения для тех, кто считает, что искру жизни способна высечь только мембрано-насосно-канальная «зажигалка». Пшибыльский и Страттен ( Przybylski,_1984 ; Stratten, 1984 ) задались интересным вопросом: а можно ли модель Ходжкина-Хаксли применить к микросферам Фокса? Они показали, что, как ни странно, нет никаких ни логических, ни физических запретов на использование этого математического аппарата для описания потенциалов действия, генерируемых этими клеточными моделями, казалось бы бесконечно далекими от нейронов. Иначе говоря, модель Ходжкина-Хаксли одинаково хорошо совместима и с нервной клеткой, и со сгустком протеиноподобных макромолекул. К сожалению, дальше этой интересной констатации Пшибыльский и Страттен не пошли. Структура модели Ходжкина-Хаксли оказалась не чувствительной (инвариантной) к физическим, химическим и структурным различиям сравниваемых объектов, вероятно, потому, что их общей субстратной основой являются белки. На первый взгляд кажется, что здесь кроется какая-то досадная ошибка или недоразумение, но в следующую минуту вспоминаешь о важнейшем преимуществе математики, как инструменте научного познания, - о ее часто непостижимой абстрактной природе. Математические понятия, уравнения, величины не обременены конкретным вещественным содержанием. У математических преобразований, на которых строятся разнообразные модели, теоремы и следствия из них, свои, особые, логические законы, а однажды полученный математический результат может иметь множество физических интерпретаций. Законы геометрии, например, не зависят от того, сделан треугольник из ржавой проволоки или образован лазерными лучами. С моделью Ходжкина-Хаксли случилось так, что ее создавали не как самодостаточный математический аппарат, а как инструмент анализа свойств предельно конкретного физического объекта - возбудимой мембраны нейрона, - и в конкретном историческом контексте! Математические параметры модели были истолкованы ее авторами, исходя из свойств аксона, но это вовсе не означает, что такая модель применима только к отростку нейрона и не допускает иных интерпретаций, когда применяется для изучения других структур. Иными словами, модель Ходжкина-Хаксли не может монополизировать живую возбудимую мембрану как единственный объект, к которому она может быть корректно применена, тем более она не может рассматриваться как математическое доказательство наличия в биомембране липидов, специфических ионных каналов и трансмембранного транспорта. Любая физическая модель - лишь одно из множества возможных воплощений духа математики и аксон здесь не исключение. Всем создателям математических моделей следует помнить, что созданная ими модель никогда не будет хранить верность той физической конструкции, с которой ее первоначально связали. Что же следует из того факта, что модели Ходжкина-Хаксли безразлично для чего ее используют, для изучения нейрона или микросферы Фокса? Мы имеем дело с двумя возможными следствиями. 1. Мембрана микросфер обладает теми же свойствами, что и мембрана нейрона: в ней тоже есть липидная мембрана, играющая роль электрического изолятора, и специализированные потенциал-зависимые ионные каналы, одни для ионов Na+ , другие для K+ . При достижении пороговой деполяризации, натриевые каналы микросферы открываются, ионы Na + устремляются в микросферу, происходит резкая деполяризация мембраны, затем открываются калиевые каналы, ну, и так далее. А дальше, для восстановления потенциала покоя, необходим ионный насос, который откачает из микросферы лишние ионы Na + и закачает в протоклетку утраченные ионы K+ . Поскольку в рассматриваемых протоклетках нет ни одного «винтика» из этого механизма, то напрашивается второе следствие. 2. Реальные события, разворачивающиеся в мембране нейрона, имеют мало общего с теорией (хорошо нам известной), которая их описывает, и поэтому такая теория нуждается в такой ревизии, чтобы между живой клеткой и клеточной моделью исчезла непроходимая, зияющая пропасть, в существовании которой мы убедились. Микросферы и нейроны построены из белков, физические свойства которых остаются неизменными на протяжении миллиардов лет, следовательно, все белковые образования должный работать на одних и тех же принципах. Теория потенциала действия нервной клетки, принятая в литературе, является лишь одной из возможных интерпретаций модели Ходжкина-Хаксли, но если это единственно возможная интерпретация, то тогда нужна другая модель. Радикальный характер этих выводов объясняет, почему Пшибыльский и Страттен их не сделали, и почему на их наблюдения не обратили никакого внимания: за истекшие 30 лет их процитировали лишь в пяти статьях люди своего круга и не в связи с парадоксальными свойствами модели Ходжкина-Хаксли. Между тем то, что принято называть ионными каналами, в микросферах есть (рис. 2, B). Сравним две представленные на рисунке записи. Очевидно, что при всех количественных различиях мы имеем дело с одним и тем же физическим явлением: электрические заряды время от времени прорываются через некую преграду (изолятор) и ионный ток то возрастает, то прекращается. Это явление наблюдается как в условиях фиксированной разности потенциалов (A), так и при разности потенциалов, генерируемых самими клеточными моделями (B). Мы видим, что некие структуры живой клетки и микросферы способны изменять свою проводимость, то увеличивая сопротивление движению зарядов, то снижая его. Что касается живых клеток, то признанным препятствием движению ионов является липидная сердцевина мембранного «сэндвича», а проницаемой ее делают белковые включения, способные выполнять разнообразные функции. Пока мы остаемся в пределах живой клетки, всё выглядит логично и стройно, но стоит перенести наше внимание на микросферы, как появляются необычные и потому интересные вопросы. Рис. 2. А. Спонтанная активность АТФ-зависимых ионных каналов клеток пшеницы, зафиксированная методом локальной фиксации потенциала (DeMarchi et al., 2011). B. Спонтанная электрическая проводимость протеиноидных микросфер Фокса, записанная методом внутриклеточного отведения потенциала (Fox, 1992). Действительно, в случае микросфер мы не можем апеллировать ни к липидной мембране, ни к ионным каналам, которые принято размещать в липидной фазе. Липидов в микросферах нет, а ионные каналы (назовем их так) есть и они должны быть еще и ион-специфичными, как того требует модель Ходжкина-Хаксли. Данные рис. 2 доказывают, что между живой клеткой и моделью нет непроходимой пропасти, что и следовало ожидать, но остается явный и глубокий разрыв в понимании природы этого сходства. Итоги Проведенное обсуждение приводит нас к следующим выводам: 1. Идея калиевых прудов представляет определенный интерес, но она не приближает нас к пониманию происхождения живой протоклетки. 2. С точки зрения существующих представлений, ключевым событием в происхождении жизни является возникновение неравновесного физического процесса, носителем которого является, как принято думать, липидная мембрана с белковыми включениями -каналами и насосами. Эта минимальная структура жизни, нуждается в непрерывном притоке энергии откуда-то извне. Значит, одновременно с полнофункциональной мембраной должна возникнуть еще и система энергообеспечения, что в целом превращает возникновение жизни в акт творения, - настолько сложные процессы синтеза и сборки должны возникнуть в одно время, в одном месте и в определенном порядке. За всё время существования проблемы происхождения жизни как экспериментальной науки ( Oparin , 1924 ), ни в одной лаборатории не было получено ни одной клеточной модели, которая обладала бы активно работающей структурой (мембраной), потребляющей энергию из внешнего источника, способной распознавать физиологически значимые катионы и направлять их движение по разным каналам, перемещая их по разные стороны первобытной мембраны. Мы до сих пор не имеем ни малейшего представления о предке современной натриевой помпы. Даже те, кто работал с липидными везикулами, со встроенной в них Na , K -АТФазой, не осмелились всерьёз предложить эти пузырьки в качестве прообраза первобытной клетки. Причина ясна: трудно представить себе, что столь сложная лабораторная технология могла реализующуюся в каком-то первобытном водоеме. 3. Общепринятые физические принципы работы клетки, которые мы встречаем повсюду, и в статьях и в учебниках, применимы только к живым клеткам и не могут, как оказалось, быть использованы для объяснения физических свойств клеточных моделей - протеиноидных микросфер. Отсутствие в литературе содержательного, предметного сопоставления физических свойств живых клеток с аналогичными свойствами их моделей наводит на странную мысль о существовании двух физик белковых тел: одна имеет дело с липидами, каналами и насосами, а другая, неизвестная пока нам физик а, управляет работой протоклеток Фокса. «Физики» почему-то разные, а явления, которые они призваны объяснять, одинаковые: потенциалы покоя и действия, канальная проводимость, способность отличать K + от Na+ . Законы физики едины и поэтому физические принципы, определяющие организацию и функционирование клетки и протоклетки, должны быть одними для всех с начала времен. Основание для этого утверждение одно: фундаментом жизни во все эпохи были белки, свойства которых зависят от их состава, а не от смены геологических периодов. Однако очевидно, что в научной среде пока нет такого понимания единства физических законов, управляющих свойствами и взаимодействиями белков, порождающими явление жизни. Тут будет, видимо, полезно ввести «принцип инвариантности физических свойств белков»: физические свойства белков не зависят от того, в состав какой структуры они входят, живой или протоклеточной. Как мы видели, модель Ходжкина-Хаксли не в состоянии дать объяснение этой инвариантности. В заключение вернемся к идее калиевых прудов. В работе Ишимы и сотр. ( Ishima et al.,_1981 ) приводятся уникальные данные о распределении ионов между протеиноидными микросферами и средой. Насколько мне известно, это единственная работа, в которой такие данные имеются. В таблице 1 этой статьи читаем: концентрация K + внутри микросферы 80 мМ, а в среде инкубации - 0,05 мМ. Получается, что концентрация K+ в микросфере в 1600 раз больше, чем в среде. Способность микросфер Фокса аккумулировать K+ обесценивает идею калиевого пруда как колыбели жизни. Пруды могли быть самыми разными по составу, а натриевый океан уже не кажется слишком враждебной средой для протоклеток, способных создавать свою внутреннюю среду без липидных мембран и Na , K -насосов. Существующие клеточные модели дают нам ясно понять, что все важные для предбиологической эволюции процессы могли протекать внутри протоклеток в относительной изоляции от окружающей среды (даже в океане), без привлечения экзотических структур и механизмов. Остается только удивляться, почему аккумулирующие способности микросфер и других клеточных моделей не исследуют самым тщательным образом. Важное значение сравнительных исследований клеток и клеточных моделей не ограничено одной только проблемой происхождения жизни. Они могли бы дать нам возможность по-новому взглянуть и на процессы в живых клетках, которые мы, якобы, хорошо понимаем. Модель Ходжкина-Хаксли - яркое свидетельство ограниченности наших знаний. Литература De Marchi U., Checchetto V., Zanetti M., Teardo E., SoccioM., Formentin E., Giacometti G. M., Pastore D., Zoratti M. and Szabò I. (2011).ATP-sensitive cation-channel in wheat (Triticum durum Desf.): identification and characterization of a plant mitochondrial channel by patch-clamp. Cellular Physiology and Biochemistry , 26 (6), 975-982. www.biophys.ru/archive/protocell/DeMarchi_2011.pdf Dubina M.V., Vyazmin S.Y., Boitsov V.M., Nikolaev E.N., Popov I.A., Kononikhin A.S., Eliseev I.E. and Natochin Y.V. (2013). Potassium ions are more effective than sodium ions in salt induced peptide formation. Origins of life and Evolution of Biospheres , 43 (2), 109-117. www.biophys.ru/archive/protocell/Dubina_2013.pdf Fox S.W. (1992). Thermal proteins in the first life and in the "mind-body" problem. In: Evolution of Information Processing Systems (pp. 203-228). Springer Berlin Heidelberg. www.biophys.ru/archive/protocell/Fox_1992.pdf Hodgkin, A. L., and Huxley, A. F. (1952). A quantitative description of membrane current and its application to conduction and excitation in nerve. The Journal of physiology , 117 (4), 500-544. www.biophys.ru/archive/protocell/Hodgkin_1952.pdf Ishima Y., Przybylski A.T. and Fox S.W. (1981). Electrical membrane phenomena in spherules from proteinoid and lecithin. BioSystems , 13 (4), 243-251. www.biophys.ru/archive/protocell/Ishima_1981.pdf Jockusch R.A., Lemoff A.S. and Williams E. R. (2001). Effect of metal ion and water coordination on the structure of a gas-phase amino acid. Journal of the American Chemical Society, 123(49), 12255-12265. www.biophys.ru/archive/protocell/Jockusch_2001.pdf Mulkidjanian A.Y. and Galperin M.Y. (2007). Physico‐Chemical and Evolutionary Constraints for the Formation and Selection of First Biopolymers: Towards the Consensus Paradigm of the Abiogenic Origin of Life. Chemistry and biodiversity , 4 (9), 2003-2015. (Статья опубликована в сентябре). www.biophys.ru/archive/protocell/Mulkidjanian_2007.pdf Mulkidjanian A.Y., Bychkov A.Y., Dibrova D.V., Galperin M.Y. and Koonin E.V. (2012). Origin of first cells at terrestrial, anoxic geothermal fields. Proceedings of the National Academy of Sciences , 109 (14), E821-E830. www.biophys.ru/archive/protocell/Mulkidjanian_2012.pdf Natochin Y.V. (2007). The physiological evolution of animals: sodium is the clue to resolving contradictions. Herald of the Russian Academy of Sciences , 77 (6), 581-591. www.biophys.ru/archive/protocell/Natochin_2007.pdf Наточин Ю.В. (2007). Физиологическая эволюция животных: натрий - ключ к разрешению противоречий. Вестник РАН, 77(11), 999-1010. (Статья опубликована в ноябре). www.biophys.ru/archive/protocell/Natochin_2007_1.pdf Oparin A.I. (1924). Proiskhozdenie Zhizny. Izd. Moskovskii Rabochii, Moscow. Przybylski A.T. (1984). Physical background of excitability: synthetic membranes and excitable cells. In Molecular Evolution and Protobiology (pp. 253-266). Springer US. www.biophys.ru/archive/protocell/Przybylski_1984.pdf Stratten W.P. (1984). Protocell action potentials: a new perspective of bio-excitation. In Molecular Evolution and Protobiology (pp. 233-251). Springer US. www.biophys.ru/archive/protocell/Stratten_1984.pdf Switek B (2012) Debate bubbles over the origin of life. Nature. doi:10.1038/nature. 2012.10024 www.biophys.ru/archive/protocell/Switek_2012.pdf
|
Дизайн и поддержка: Interface Ltd. |
|